
The Little Rouge River in March 2020 (photo by Nina Munteanu)

Caved-in path along Little Rouge (photo N. Munteanu)
It is early spring in the Little Rouge River woodland and the river that had swollen with snow melt just a week before, now flows with more restraint. I can see the cobbles and clay of scoured banks under the water. Further on, part of the path along the river has collapsed from a major bank scour the previous week. The little river is rather big and capricious, I ponder; then I consider that the entire forest sways to similar vagaries of wind, season, precipitation and unforeseen events. Despite its steadfast appearance, the forest flows—like the river—in a constant state of flux and change, cycling irrevocably through life and death.
The forest appears caught between the slumber of winter and the vigorous awakening of spring. It is the time of the spring equinox. Like a halting breath, the forest is poised in a hush. A quietness, a kind of hesitation—as with dawn or dusk—inhabits the forest.

Little Rouge swells over its banks, then a week later settles (photos by Nina Munteanu)
I walk slowly, eyes cast to the forest floor to the thick layer of dead leaves, and discover seeds and nuts—the promise of new life. I aim my gaze past trees and shrubs to the nearby snags and fallen logs. I’m looking for hidden gifts. One fallen cedar log reveals swirling impressionistic patterns of wood grain, dusted with moss and lichen.

Cedar log with wood patterns from erosion and decay (photo by Nina Munteanu)

Beetle bore holes drilled into Pine bark (photo by Nina Munteanu)
The bark of a large pine tree that has fallen is riddled with tiny beetle holes drilled into its bark. Where the bark has sloughed off, a gallery of larval tracks in the sapwood create a map of meandering texture, form and colour.
Nearby, another giant pine stands tall in the forest. Its roughly chiselled bark is dusted in lichens, moss and fungus. The broad thick ridges of the bark seem arranged like in a jigsaw puzzle with scales that resemble metal plates. They form a colourful layered mosaic of copper to gray and greenish-gray. At the base of the tree, I notice that some critter has burrowed a home in a notch between two of the pine’s feet.

Beetle larva tracks (gallery) in Pine wood (photo by N. Munteanu)
Then just around the corner, at the base of a cedar, I spot several half-eaten black walnuts strewn in a pile—no doubt brought and left there by some hungry and industrious squirrel who prefers to dine here.
The forest ecosystem supports a diverse community of organisms in various stages of life and death and decay.

Half-eaten black walnuts strewn at base of Cedar tree (photo N. Munteanu)
Trees lie at the heart of this ecosystem, supporting a complex and dynamic cycle of evolving life.
Even in death, the trees continue to support thriving detrivore and saprophytic communities that, in turn, provide nutrients and soil for the next generation of living trees. It’s a partnership.
Decomposition and decay are the yin to the yang of growth, writes Trees for Life; and together they form two halves of the whole that is the closed-loop cycle of natural ecosystems.

Cedar snags with woodpecker holes (photo by Nina Munteanu)

Cedar snag with large Woodpecker hole (photo by Nina Munteanu)
The forest is littered with snags and fallen trees in different stages of breakdown, decomposition and decay. I spot several large cedar, pine, oak and maple snags with woodpecker holes. Tree snags may remain for many decades before finally falling to the ground.
Snags provide cavity nest sites, platforms and feeding substrates for raptors, other birds and small mammals. They also serve as overwintering sites, lookout posts and roosting sites for many wildlife. Snags and rotting logs on the forest floor provide damp shelter and food for many plants and animals. Most are decomposers, including earthworms, fungi, and bacteria. As the wood decays, nutrients in the log break down and recycle in the forest ecosystem. Insects, mosses, lichens, and ferns recycle the nutrients and put them back into the soil for other forest plants to use. Dead wood is an important reservoir of organic matter in forests and a source of soil formation. Decaying and dead wood host diverse communities of bacteria and fungi.
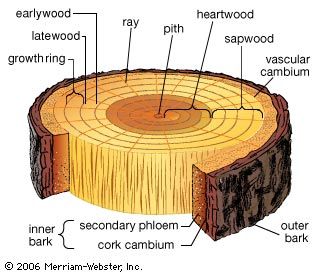 Wood tissues of tree stems include the outer bark, cork cambium, inner bark (phloem), vascular cambium, outer xylem (living sapwood), and the inner xylem (non-living heartwood). The outer bark provides a non-living barrier between the inner tree and harmful factors in the environment, such as fire, insects, and diseases. The cork cambium (phellogen) produces bark cells. The vascular cambium produces both the phloem cells (principal food-conducting tissue) and xylem cells of the sapwood (the main water storage and conducting tissue) and heartwood.
Wood tissues of tree stems include the outer bark, cork cambium, inner bark (phloem), vascular cambium, outer xylem (living sapwood), and the inner xylem (non-living heartwood). The outer bark provides a non-living barrier between the inner tree and harmful factors in the environment, such as fire, insects, and diseases. The cork cambium (phellogen) produces bark cells. The vascular cambium produces both the phloem cells (principal food-conducting tissue) and xylem cells of the sapwood (the main water storage and conducting tissue) and heartwood.

Uprooted stump covered in carbon cushion fungi, moss and lichen (photo by Nina Munteanu)
Tree Decay

Wood decay from insect boring and black rot fungi (photo by Nina Munteanu)
When a tree falls, it eventually starts to decompose, creating unique new habitats in its body as the outer and inner bark, sapwood, and heartwood rot at different rates. Decomposition increases the flow of plant and animal populations, air, water, and nutrients between a fallen tree and its surroundings. Once organisms gain entrance to the interior, they consume and break down wood cells and fibers. Larger organisms-mites, collembolans, spiders, millipeds, centipedes, amphibians, and small mammals patiently await the creation of internal spaces before they can enter. Wood-rotting fungi decay spring wood faster than summer wood, creating zones of weakness, especially between annual rings.
The tree’s different woody tissues show different susceptibilities to decay, which normally begins at the exterior of a tree and progresses inward. The outer bark disappears mainly by fragmentation and sloughing from the top and sides of fallen trees. Despite its high nitrogen content, decay of outer bark is inhibited by its high lignin content and other anti- microbial and nutrients. The outer ring of living sapwood is usually highly resistant to decay despite its high nutrient content, and may remain so even when adjacent tissues are extensively decayed. Dead sapwood decays rapidly, however. Compared to sapwood, heartwood decays slowly because of low nutrient quality and digestibility to microbes (high lignin and low nitrogen).

Forest ecologists define five broad stages in tree decay, shown by the condition of the bark and wood and presence of insects and other animals. The first two stages evolve rapidly; much more time elapses in the later stages, when the tree sags to the ground. These latter stages can take decades for the tree to break down completely and surrender all of itself back to the forest. A fallen tree nurtures, much like a “mother” archetype; it provides food, shelter, and protection to a vast community—from bears and small mammals to salamanders, invertebrates, fungus, moss and lichens. This is why fallen trees are called “nursing logs.”

Maple log in various stages of decay (photo by Nina Munteanu)
I stop to inspect another fallen tree lying on a bed of decaying maple, beech and oak leaves. When a fallen tree decomposes, unique new habitats are created within its body as the outer and inner bark, sapwood, and heartwood decompose at different rates, based in part on their characteristics for fine dining. For instance, the outer layers of the tree are rich in protein; inner layers are high in carbohydrates.
This log—probably a sugar maple judging from the bark that remains—has surrendered itself with the help of detrivores and saprophytes to decomposition and decay. The outer bark has mostly rotted and fallen away revealing an inner sapwood layer rich in varied colours, textures and incredible patterns—mostly from fungal infestations.

Rotting maple covered in carbon cushion and green-stain fungi, moss and lichen (photo by Nina Munteanu)
In fact, this tree is a rich ecosystem for dozens of organisms. Wood-boring beetle larvae tunnel through the bark and wood, building their chambers and inoculating the tree with microbes. They open the tree to colonization by other microbes and small invertebrates. Slime molds, lichen, moss and fungi join in. The march of decay follows a succession of steps. Even fungi are followed by yet other fungi in the process as one form creates the right condition for another form.
Most hardwoods take several decades to decompose and surrender all of themselves back to the forest. In western Canada in the westcoast old growth forest, trees like cedars can take over a hundred years to decay once they’re down.

Fragmented sapwood with spalting from fungus (photo by Nina Munteanu)
The maple log I’m studying in this Carolinian forest looks like it’s been lying on the ground for a while, certainly several years. The bark has fragmented and mostly fallen away, revealing layers of sapwood and heartwood in differing stages of infestation and decay. Some sapwood is fragmented and cracked into blocks and in places looks like stacked bones. Much of the heartwood—the interior core of the tree trunk made of cellulose and lignin—is rotting in columns and turning into sawdust.
Spalting & Intarsia
Black lines as though drawn by a child’s paintbrush flow through much of the sapwood; these winding thick streaks of black known as “zone lines” are in fact clumps of dark mycelia, which cause “spalting,” the colouration of wood by fungus. According to mycologist Jens Petersen, these zone lines prevent “a hostile takeover by mycelia” from any interloping fungi. Most common trees that experience spalting include birch, maple, and beech. Two common fungi that cause spalting have colonized my maple log. They’re both carbon cushion fungi.

Spalting by carbon cushion fungus (photo by Nina Munteanu)
Brittle cinder (Kretzschmaria deusta) resembles burnt wood at maturity. Deusta means “burned up” referring to the charred appearance of the fungus. Hypoxylon forms a “velvety” grey-greenish cushion or mat (stroma) that resembles hardening pahoehoe lava. As the Hypoxylon ages, it blackens and hardens and tiny, embedded fruitbodies (perithecia) show up like pimples over the surface of the crust.

Hypoxylon, showing “pimples” and fruiting bodies (photo by Nina Munteanu)
Much of the exposed outer wood layer looks as though it has been spray painted with a green to blue-black layer. The “paint” is caused by the green-stain fungus (Chlorociboria) and blue-stain fungus (Ceratocystis). The blue-green stain is a metabolite called xylindein. Chlorociboria and Ceratocystis, also spalter fungi, spread throughout the wood in search of sugars, colonizing the ray cells first then the vessels and fibres as they release the xylindein metabolite and change the colour of the wood. According to woodworkers, the dye does not oxidize or fade–showing colorfastness–and apparently a mycological rarity.

Various stain and black cushion fungi on decaying maple log (photo by Nina Munteanu)

Intarsia woodwork using spalting fungus
While zone lines that create spalting don’t damage wood, the fungus responsible most likely does.
Spalting is common because of the way fungi colonize, in waves of primary and secondary colonizers.
Primary colonizers initially capture and control the resource, change the pH and structure of the wood, then must defend against the secondary colonizers now able to colonize the changed wood.
Wood that is stained green, blue or blue-green by spalting fungi has been and continues to be valued for inlaid woodwork called intarsia. In an article called “Exquisite Rot: Spalted Wood and the Lost Art of Intarsia” Daniel Elkind writes of how “the technique of intarsia–the fitting together of pieces of intricately cut wood to make often complex images–has produced some of the most awe-inspiring pieces of Renaissance craftsmanship.” The article explores “the history of this masterful art, and how an added dash of colour arose from the most unlikely source: lumber ridden with fungus.”

Blue and green-stain fungi join others in spalting wood (photo by Nina Munteanu)
Moss

Moss joins lichen in colonizing branch of a shrub in Little Rouge woodland (photo by Nina Munteanu)
I find moss everywhere in the Little Rouge woodland, including beneath the forest floor. Moss is ubiquitous, adapting itself to different situations and scenarios.

Moss growing up a cedar tree (photo by Nina Munteanu)
Like a shapeshifter, moss is at once coy, hiding beneath rotting leaf litter, stealthy and curious as it creeps up the feet of huge cedars, and exuberant as it unabashedly drapes itself over every possible surface such as logs, twigs and rocks, and then proceeds to procreate for all to see.
Moss is a non-vascular plant that helps create soil; moss also filters and retains water, stabilizes the ground and removes CO2 from the atmosphere. Science tells us that mosses are important regulators of soil hydroclimate and nutrient cycling in forests, particularly in boreal ecosystems, bolstering their resilience. Mosses help with nutrient cycling because they can fix nitrogen from the air, making it available to other plants.

Moss gametophytes with sporophytes growing on them (photo by Nina Munteanu)
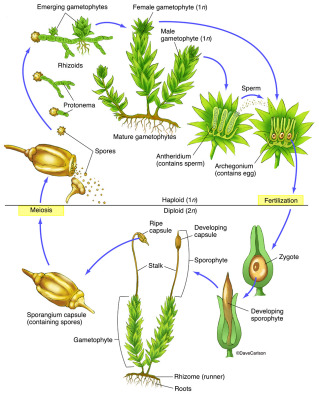 Mosses thrive in the wet winter and spring, providing brilliant green to an otherwise brown-gray environment. Even when covered in snow (or a bed of leaves), moss continues its growth cycle, usually in the leafy gametophyte stage. When the winter is moderate, like it is near Toronto, sporophyte structures can already appear on stalks that hold a capsule full of spores. In the spring the capsules release spores that can each create a new moss individual.
Mosses thrive in the wet winter and spring, providing brilliant green to an otherwise brown-gray environment. Even when covered in snow (or a bed of leaves), moss continues its growth cycle, usually in the leafy gametophyte stage. When the winter is moderate, like it is near Toronto, sporophyte structures can already appear on stalks that hold a capsule full of spores. In the spring the capsules release spores that can each create a new moss individual.
The dominant phase of the moss life-cycle is the gametophyte (haploid) phase—the typical green moss plant that most of us see. The gametophyte consists of rhizoids that attach it to a substrate, a simple or branched stem, and small leaves (mostly one cell thick).

Moss on a log with maturing sporophytes (photo by Nina Munteanu)
When there is enough moisture, the male and female gametes produce the short-lived second phase, the sporophyte. The sporophyte “foot” embeds into the gametophyte and forms a stalk and capsule filled with spores. When the spores mature, the stalk bends and a tiny lid opens to release the diploid spores in the wind. Mosses also reproduce asexually (by groups of cells called gemmae or deciduous shoots called bulbils) or by fragmentation (e.g. breaking off almost any plant part, which then grows into a new plant).
In boreal forests, some moss provide nitrogen for the ecosystem through their relationship with nitrogen-fixing cyanobacteria (blue-green algae). The cyanobacteria colonizes moss and receives shelter in return for providing fixed nitrogen. Moss releases the fixed nitrogen, along with other nutrients, into the soil of the ecosystem when disturbed by drying-rewetting and fire events.
Moss is quietly, gloriously profligate.
Lichen

Physchia stellaris lichen and others colonizing twig on forest floor (photo by Nina Munteanu)

close up of the apothecia of Physchia (photo by Nina Munteanu)
Many twigs strewn on the leaf-covered forest floor were covered in grey-green lichen with leaf-like, lobes. On close inspection, the lichen thallus contained abundant cup-shaped fruiting bodies. These lichen were most likely Physchia stellaris, common and widespread in Ontario and typically pioneers on the bark of twigs, particularly of poplars, and alders.
 Lichens are a complex symbiotic association of two or more fungi and algae (some also partner up with a yeast). The algae in lichens (called phycobiont or photobiont) photosynthesize and the fungus (mycobiont) provides protection for the photobiont. Both the algae and fungus absorb water, minerals, and pollutants from the air, through rain and dust. In sexual reproduction, the mycobiont produces fruiting bodies, often cup-shaped, called apothecia that release ascospores. The spores must find a compatible photobiont to create a lichen. They depend on each other for resources—from food to shelter and protection.
Lichens are a complex symbiotic association of two or more fungi and algae (some also partner up with a yeast). The algae in lichens (called phycobiont or photobiont) photosynthesize and the fungus (mycobiont) provides protection for the photobiont. Both the algae and fungus absorb water, minerals, and pollutants from the air, through rain and dust. In sexual reproduction, the mycobiont produces fruiting bodies, often cup-shaped, called apothecia that release ascospores. The spores must find a compatible photobiont to create a lichen. They depend on each other for resources—from food to shelter and protection.
 Lichens also reproduce asexually in two ways. One way is through fragmentation; a portion of the thallus simply breaks off and starts a new colony. The second way is through vegetative structures called soredia—algal cells surrounded by fungal filaments or hyphae—which form in the thallus. In the right conditions, they burst through the surface of the thallus and as propagules (such as a stem cuttings, portions of root, seeds or spores) can propagate more of themselves. A lichen might also develop a specific vegetative extension called isidia that fragments as a propagule to create more lichen.
Lichens also reproduce asexually in two ways. One way is through fragmentation; a portion of the thallus simply breaks off and starts a new colony. The second way is through vegetative structures called soredia—algal cells surrounded by fungal filaments or hyphae—which form in the thallus. In the right conditions, they burst through the surface of the thallus and as propagules (such as a stem cuttings, portions of root, seeds or spores) can propagate more of themselves. A lichen might also develop a specific vegetative extension called isidia that fragments as a propagule to create more lichen.
Turkey Tail Fungus
Turkey Tail Mushrooms are one of the most common mushrooms in the forest, and they grow all around the world. They grow on almost any type of wood, and there are no known poisonous look-alikes.

Turkey Tail fungus on tree in Little Rouge woodland (photo by Nina Munteanu)
Turkey Tail bracket fungus (Trametes versicolor / Coriolus versicolor) is a white rot fungus that breaks down the lignin and to some degree the cellulose and hemi-cellulose in wood. The mushroom resembles a turkey tail and is a polypore. This allows it to be distinguished from a similar looking and equally common mushroom, the False Turkey Tail (Stereum hirsutum), a crust fungus.
The best way to check, according to mycologist Debbie Viess is to look at its underside. “The underside of a mushroom is called the hymenium or fertile surface, where spores are produced and then drop to the ground. As a polypore, Turkey Tail holds its spores in tubes, so its underside should display tiny holes visible to the naked eye.” The pores are just visible, about 3-8 pores per millimeter. False Turkey Tail, which also has colourful rings on the top surface, has no pores on its underside, which is completely smooth.
Another similar looking mushroom is Trichaptum abietinum, also a polypore. “But rather than pores or a smooth surface,” says Viess, “it has little teeth or bumps of tissue where its spores are produced. Plus, you can’t miss its brilliant lilac cast.”

Left to right: Trametes versicolor, Stereum hirsutum, and Trichaptum abietinum
Turkey Tail is well known for its immune-boosting benefits and additional medicinal properties, particularly its cancer preventative properties. Packed with anti-oxidants such as phenols and flavonoids, this fungus is not palatable due to its tough texture. When fresh it is best used as a tea. Collecting turkey tails and drying them out is recommended. Once dried they can still be used to make tea, to add to other teas, and used in soups and in other meals.
I haven’t tried it yet, but I certainly intend to. Foraging here I come!

Black Witches Butter (Exidia glandulosa) colonizes twig on forest floor of The Little Rouge (photo by Nina Munteanu)

Nina Munteanu is a Canadian ecologist / limnologist and novelist. She is co-editor of Europa SF and currently teaches writing courses at George Brown College and the University of Toronto. Visit www.ninamunteanu.ca for the latest on her books. Nina’s bilingual “La natura dell’acqua / The Way of Water” was published by Mincione Edizioni in Rome. Her non-fiction book “Water Is…” by Pixl Press (Vancouver) was selected by Margaret Atwood in the New York Times ‘Year in Reading’ and was chosen as the 2017 Summer Read by Water Canada. Her novel “A Diary in the Age of Water” will be released by Inanna Publications (Toronto) in May 2020.

2 thoughts on “The Hidden Forest: Gems of the Little Rouge Woodland”